
Les Algues sont présentes dans le monde entier et se développent dans toutes les étendues d’eau ou dans tout milieu terrestre où le taux d’humidité reste élevé. Elles vivent aussi en symbiose avec d’autres organismes. Certaines espèces peuvent même survivre sur la neige ou la glace des régions polaires ou montagneuses, ou supporter les températures élevées des sources d’eau chaude.
Les Algues sont des thallophytes autotrophes photosynthétiques d’organisation simple, formant un groupe extrêmement hétérogène du point de vue de leur origine évolutive. Pour cette raison, leur classification controversée est d’une complexité rare.
I- Evolution et phylogénie.
A- Que sont les algues ?
On ne peut pas donner d’image typique des algues. Les couleurs observées, bien que différentes, ne sont pas spécifiques.
On trouve des algues de quelques micromètres (diatomées) jusqu’à plusieurs mètres. à On a une grande diversité : diversité de taille ; diversité morphologique : les Chlamydomonas sont des unicellulaires, Ectocarpus est filamenteuse, Ulva lactuca est structurée ; diversité écologique : on peut trouver des thallophytes dans les cours d’eau (diatomées, algues vertes, cyanobactéries), dans la neige (algues vertes) et sur terre.
Les laminaires présentent une convergence avec les végétaux supérieurs.
==> Finalement, on observe de fortes adaptations aux différents milieux.
B- Position phylogénétique : systématique et définitions.
Les thallophytes regroupent tout ce qui est différent des cormophytes.
Chez ces thallophytes, une cellule mère va générer des cellules filles emprisonnées dans la paroi de cette cellule mère.
Chez Chara, on trouve des cellules de reproduction sexuée qui préfigurent la graine. Les Cryptogames ont des structures de reproduction cachées alors que les Phanérogames ont des structures de reproduction visibles.
Les caractères utilisés en phylogénie sont :
- Les caractères ontogéniques : le passage par des phases archaïques.
- Les caractères paléontologiques.
- La morphologie comparée.
- Les caractères moléculaires : ces caractères peuvent être discontinus, continus. On prendra le cas des gènes 16s. Ce dernier est très utilisé pour la mise en place des phylogénies car il est présent chez tous les organismes. Ils sont aussi abondants et très faciles à isoler.
Les algues sont des eucaryotes autotrophes dont l’appareil végétatif est un thalle (sans tissu différencié). On leur rapproche les cyanobactéries et les prochlorobactéries.
1- Ectocarpus siliculosus.
L’appareil reproducteur est un cyste pluriloculaire avec des grains formés par mitoses. Lorsque l’on n’a qu’une loge, c’est un cyste uniloculaire, porté par des cellules contenant plusieurs spores.
Quand on n’a pas de cyste uniloculaire, on se trouve chez le gamétophyte.
2- Sphacelaria plumula.
On trouve une grosse cellule apicale et une tige formée d’une seule rangée de cellules.
Autour de l’axe central, on a un cortex cellulaire.
Les ronds orangés sont des plastes.
==> Le thalle est uniaxial, polystylé.
3- Fucus.
Fucus est une espèce dioïque. Les points observés à la surface sont des conceptacles. Les très grosses granules sont les parties femelles. Les tas de filaments sans structure épaisse sont la partie mâle.
4- Laminaria digitata.
On trouve deux zones corticales pigmentées (le méristoderme) avec des structures de reproduction (cystes et paraphyses). On trouve ensuite deux zones composées de grosses cellules (les cellules d’assises) et une zone interne avec des filaments (la médullaire).
On trouve un plaste divisé en vésicules, reliées entre elles par une membrane.
II- Diversité de thalle et mécanisme de croissance.
A- Organisation générale.
Les archéthalles présentent des cellules non unies (indépendantes), isolées ou non.
Les nématothalles montrent des cellules dépendantes avec des thalles filamenteux pouvant évoluer en lame. La croissance va être diffuse ou localisée en région apicale. Les cellules sont reliées par le plasmodesme.
Les cladothalles sont des thalles filamenteux à axe de croissance préférentiel.
1- Les Archéthalles.
a- Les cellules les plus simples.
On trouve des cellules indépendantes, isolées ou incluses dans un cénobe. On trouve par exemple, Chlamydomonas qui possède deux flagelles.
b\ Les cellules isolées.
Dans ces cellules, on trouve : une paroi, un corps intracellulaire, des dyctiosomes, un gros plaste avec une masse protéique (pyrénoïde), un stigma dans le plaste (la structure photosensible qui permet l’orientation des cellules nageuses), deux vacuoles pulsatiles participant à l’osmorégulation, deux équiflagelles. La division a généralement lieu par bipartition.
c- Les associations de cellules immobiles.
Chez Scénédesmus, on a quatre cellules qui peuvent se séparer et donner de nouveaux cénobes. Ces cellules sont associées par un ciment.
d- Les thalles pluricellulaires.
Dans ce cas, on peut trouver des cyanobactéries a structure pluricellulaire.
- Chez Zygnéma, les cellules sont indépendantes, non réunies par un plasmodème.
- Chez Pediastrum, il y a association cellulaire. Chaque cellule donnera un cyste qui permettra la formation de spores flagellées restant groupées dans un mucilage. L’appareil végétatif va se transformer et donner un adulte : les centres opérateurs organisent la mise en place du réseau de filaments.
e- Les associations cellulaires mobiles.
- Chez les chlorophycées, on observe une évolution de ces structures.
De Gonium, groupement de 16 cellules en forme de coupelle, on passe à Pandorina, comprenant toujours 16 cellules mais cette fois, de forme sphérique. Finalement, on arrive à Eudorina, composée de 64 cellules avec une spécialisation apparente. Le dernier stade est Volvox.
- Chez Volvox, composé de plusieurs milliers de cellules, on trouve des petits ronds verts (à l’intérieur) qui sont des cénobes fils.
Cette évolution morphologique va être suivie par une spécialisation des cellules (chez Gonium, toutes les cellules sont indépendantes). Chez Eudorina, de nombreuses cellules ont une fonction reproductrice. La tendance évolutive est à la réduction du nombre de cellules reproductrices.
- Chez Volvox, les gonidies (appareils reproducteurs) apparaissent au moment de la reproduction. Ces gonidies sont des retournements de membrane.
2- Les Nématothalles.
a- Origine.
- Chez les nématothalles, on observe un changement d’organisation fonctionnelle par rapport aux archéthalles.
- Ces thalles peuvent évoluer en lame.
- Les relations intercellulaires se font par l’intermédiaire d’une synapse, bouchée par un bouchon synaptique
b- Les divers types de filaments.
- On peut trouver des filaments prostrés, dressés, hétérotriches.
- Les filaments haplostiques sont des filaments composées d’une unique file cellulaire (exemple : Ectocarpus).
- Les filaments polystiques sont composés de plusieurs rangées de cellules (exemple : Entéromorpha).
- Parmi les espèces à thalle siphonné, on trouve Vaucheria.
3- Les cladothalles.
a- Origine.
Ces thalles ont une axe préférentiel de croissance : le cladome primaire. La croissance est continue. Sur chaque cladome, on va trouver des pleuridies. Les pleuridies sont de petits filaments ayant une croissance continue.
b- Les différentes structures.
- Les cladothalles uniaxiaux ont un axe central composé d’une file de cellules (haplostiques).
- Les cladothalles polystiques ont un axe central avec cortex.
- Les cladothalles multiaxiaux présentent des articulations entre des articles calcifiés : c’est une zone sous le cortex pleuridien (exemple : Corallina officinalis).
c- Variations morphologiques.
- Les lames foliacées (exemple : Delesseria) : La lame donne un cladome primaire qui donne lui, des cladomes secondaires et des pleuridies couchées entre les cladomes secondaires.
- Les cladomes à filaments siphonnés : Ils sont trouvés chez les Choloropyceae, Bryopsidales.
- Les cladomes à filaments hémisiphonés: Dans ce cas, on a des articles plurinucléés. Entre les cellules, la paroi vient former un bouchon d’obturation. Des vacuoles internes, riches en polysaccharides, viennent former la paroi.
- Les cladomes de Florideophyceae : Ces cladomes présentent de nombreuses variantes ; ils peuvent être formés de cellules ou de filaments ==> ce sont des cladomes aplatis.
On trouve aussi des cladomes multiaxiaux : une zone corticale est formée de filaments ou de cellules ainsi que de petits rameaux. Chez certaines algues, le cladome multiaxial disparaît sauf dans la région apicale.
B- Mode de croissance.
1– La croissance diffuse.
Toutes les cellules sont aptes à se diviser : Chlorophyceae, Ulothrix, Spyrogyra.
2- La croissance localisée (une zone de croissance).
On peut trouver une zone intercalaire qui est contenue dans l’appareil végétatif.
La croissance trichothallique se fait par des cellules supérieures plus fines et elle est rencontrée dans plusieurs genres d’algues brunes comme chez Ectocarpus. On peut aussi trouver plusieurs zones de croissance par filament.
3- La croissance apicale.
Cette croissance est réalisée par une cellule apicale et permet la formation de toutes les algues siphonnées. On trouve ce cas chez les Sphacélariales (algues brunes). Si l’on ne trouve qu’une cellule apicale, on aura un cladome uniaxial ; si l’on a plusieurs cellules apicales, le cladome sera multiaxial.
C- Les thalles de phaeophyceae.
L’appareil végétatif est très développé.
Chez Dyctiota dichotomia, on a une cellule apicale qui donne deux cellules filles puis deux ramifications qui donneront quatre cellules filles puis quatre ramifications.
Les cellules à la base des cellules apicales sont des cellules filles.
La zone corticale donne des cellules pigmentées. Les grosses cellules sont des cellules de stockage.
Fucus sp :
L’appareil végétatif est un disque basal et est aplati. Il peut donner des vésicules aérifères (pour le soutien).
- On ne trouve pas de nervure mais une rainure et une zone fertile apicale (le réceptacle).
- A l’extrémité, on trouve des cryptes avec une grosse cellule (cellules apicales) avec cinq plans de mitose. Cette cellule est l’origine d’une couche cellulaire qui sera à l’origine de filaments.
- On trouve aussi deux zones corticales (méristoderme) assimilatrices.
- On distingue aussi des cellules anguleuses qui forment un cortex de soutien.
Les Laminariales:
Les laminariales ont une longue structure érigée : stype avec une fronde et crampon basal.
On distinguera deux organisations différentes : un stype pérenne et une fronde annuelle. La zone de croissance est intercalaire et l’allongement du stype entraîne la formation d’une nouvelle fronde.
Sur le stype, à l’extérieur, on a le méristoderme qui montre une croissance en épaisseur montrée par des stries annuelles.
On peut voir des petits canaux qui sont des canaux mucifères et des cellules accolées à paroi criblée.
==> Cette ligne des laminariales est parallèle à celle qui a donné les végétaux supérieurs.
D- Les structures cellulaires particulières.
1- La paroi ou exosquelette.
a- Les algues sans paroi.
Ces algues ont un système de rigidification.
- Les Cryptophytes (unicellulaires). Il y a formation de plaques qui sont des inclusions de protéines sous le plasmalemme.
- Les Dinophyceae ont des armures de plaques protéiques.
- Les Euglenophyceae. Chez Phacus, une espèce striée, on peut voir que les stries sont dues à des plaques articulées entre elles sous le plasmalemme. Les Phacus sont capables d’effectuer des mouvements amiboïdes.
- Les Chrysophyceae (Emiliana luxleyi). A l’intérieur, on trouvera des écailles de calcite aussi appelées coccolites ou coccosphères.
Chez les Diatomophyceae (Bacillariophyceae), on trouve un frustule siliceux. Ce sont des espèces souvent pélagiques. On trouve deux types de symétrie : radiale (centrique) ou planaire (pennées). L’extérieur est recouvert par deux valves de silice, emboîtées (l’épivalve est supérieure et l’hypovalve est inférieure). Des ceintures connectives vont sceller les valves.
La division se fait quand une ceinture connective glisse ; il y aura division cellulaire et formation de nouvelles valves. Les nouvelles valves seront toujours des hypovalves (par rapport à celles déjà existantes). è Il y a donc une diminution de la taille. Plus la taille diminue, plus la flottabilité régresse. Au bout d’un moment, il va y avoir reproduction sexuée par zoospores, fusion, formation d’un embryon et formation d’une cellule mère.
b- La paroi des algues.
Cette paroi est constituée de deux phases : une phase cristalline (squelette de microfibrilles) et une phase amorphe (matrice prédominante).
Les algues rouges et brunes on une phase amorphe importante qui est un réseau de polysaccharides qui forme un mucilage (origine en boite à œufs).
La phase cristalline : Cette phase est composée de cellulose, de xylème, de mannane, avec selon les algues, un polymère dominant.
- Chez les chlorophycées, la phase amorphe est majoritairement composée d’hétéropolysaccharides sulfatés.
- Chez les Rhodophyta, on trouve des galactanes, pour résister à l’immersion.
- Chez les Phaeophyceae, on trouvera principalement des alginates.
2- Les flagelles.
Un flagelle est composé de 9 associations de tubulines (a et b) avec 250 protéines associées.
On trouve un corpuscule basal qui permet l’insertion du flagelle ainsi qu’une structure intracellulaire.
Comme organisation de l’appareil cinétique on trouve :
- Cellules acrokontées : les flagelles apicaux.
- Cellules isokontées : les flagelles sont égaux.
- Cellules pleurokontées : les flagelles sont latéraux.
- Cellules hétérokontées : les flagelles sont différents.
- Cellules stéphanokontées : les flagelles ont une disposition rayonnante.
Chez les Rhodophyta, les flagelles sont absents ; chez les Chlorophyceae, les flagelles sont isokontés ; les Hétéroconta ont des flagelles hétérokontés. Le déplacement des cellules flagellées se fait vers le haut (la fuite, vers le bas).
3-Les plastes.
a- Organisation.
Les formes archéoplastidiées : on trouve un gros plaste focal ou pariétal.
Les formes néoplastidiées : ce sont souvent des structures cladomiennes.
Les formes mésoplastidiées : ce sont des structures intermédiaires (exemple : Chladophora).
Les algues rouges, Bangyophyceae sont archéoplastidiées ; les Florideophyceae sont néoplastidiées.
Les algues brunes, comme Ectocarpus, sont archéoplastidiées.
b- Structure.
Les algues vertes ont des chloroplastes qui sont formés de deux membranes et un système interne, les thylacoïdes. Chez les végétaux supérieurs, on a des granas qui sont des empilements de thylacoïdes.
Ces algues comprennent un pyrénoïde (structure protéique interne) qui ne forme pas de grana et qui permet la synthèse d’amidon intra-plastidiale.
Les algues brunes ont des phaeoplastes. Certains plastes comprennent un pyrénoïde en position déjetée. L’enveloppe externe est formée de quatre membranes : deux membranes comme chez les chloroplastes et deux externe de réticulum endoplasmique.
Entre les deux membranes externes et les deux internes, on trouve une structure avec de l’ADN : c’est le nucléomorphe (un noyau atrophié). Il n’y a jamais de phycobilisomes.
Chez les Dinophyceae, on trouve une enveloppe classique, composée de deux membranes avec une membrane de réticulum.
Souvent, les thylacoïdes sont groupés par trois (en général). Chez les Cryptophyceae, les thylacoïdes sont groupés par 2. Ceux de Dinophyceae et Euglenophyceae sont souvent non groupés (parfois, sont par trois).
Les rhodoplastes de Rhodophyceae.
Ce sont des pigments rouges hydrosolubles. L’enveloppe plastidiale est formée de deux membranes. Souvent, on a un seul thylacoïde qui peut faire tout le tour du plaste. Il y a absence de pyrénoïde dans les thylacoïdes des algues rouges.
En microscopie électronique, on voit des points de phycobilisomes : se sont des structures protéiques qui recouvrent les photosystèmes sur les thylacoïdes. Ils contiennent de la phyco-érythrine ainsi que de la phycocyanine.
On retrouve ce système sur les cyanobactéries où les thylacoïdes sont chapeautées par des phycobilisomes.
Les Cryptophyceae présente un nucléomorphe avec quatre membranes.
Les algues rouges ont deux membranes.
c- Origine évolutive, la théorie de l’endosymbiose.
On trouve une structure unicellulaire eucaryote qui héberge un procaryote symbiotique : ce sont des cyanobactéries dans des glaucocystophytes.
Le symbiote est intégré à la cellule et subit une perte de son système membranaire (algues rouges).
Une cellule eucaryote « avale » une cellule eucaryote endosymbiotique : ce sont les cryptophytes (quatre membranes et un nucléomorphe).
Il y aura une disparition du nucléomorphe : il reste une enveloppe, deux membranes, deux réticulum mais plus de nucléomorphe.
On trouve des plastes primaires (chlorophyceae, rhodophyceae) et des plastes secondaires (algues brunes).
d- Les stigmas.
Les stigmas sont présents chez de nombreuses algues flagellées.
Un stigma est une structure composée de caroténoïdes qui selon les espèces est une dépendance d’un plaste ou un plaste dégénéré.
C’est un photosystème qui intervient dans l’orientation : c’est un cache qui permet de générer une zone d’ombre. De plus, une structure photosensible serait à la base du flagelle. La flavine est la vraie structure photosensible réceptive. On observera une variation de dépolarisation du flagelle qui le fait battre ou non.
III- La reproduction.
A- Multiplication végétative.
1- La bipartition cellulaire.
Les cellules vont se diviser en deux (exemple : Chlamydomonas). La division donne des cellules filles qui assureront la dissémination ou une réunification dans une colonie (exemple : Diatomées).
Cette bipartition n’existe pas chez les algues rouges.
2- La fragmentation du thalle.
Cette fragmentation est fréquente, même chez les pluricellulaires. Celle-ci permet au végétal d’avoir un côté invasif. On trouve par exemple Caulerpa taxifolia qui envahie les posidonies (Posidonia et Zostera sont des graminées marines). Cette caulerpe est toxique pour de nombreux poissons et coquillages et vont donc diminuer la qualité du biotope.
Par exemple, la Sargasse est une algue ayant envahie la côte européenne venue avec des huîtres du Japon.
3- La différenciation de spores.
L’exemple des Sphacélariales :
On trouvera par exemple des propagules qui sont des structures pluricellulaires.
Il pourra y avoir formation de zoospores comme chez Oedogonium (libération de spores stéphanokontés).
B- La reproduction sexuée.
1- Les gamètes et types de gamies.
a- L’hologamie.
Il va y avoir de deux cellules correspondant à l’appareil végétatif qui fusionnent. C’est le cas des structures unicellulaires.
b- La cystogamie.
Il va y avoir fusion de deux cellules sans qu’il n’y ait de différenciation de gamètes (une cellule par individu).
c- La planogamie.
Il va y avoir de gamètes flagellés. Gamètes mâles et femelles peuvent avoir la même morphologie (planogamie isogame) ou une morphologie différente (planogamie anisogame).
d- L’oogamie.
Dans ce cas, l’oosphère est non flagellée.
e- Exemples.
La cystogamie chez Casmarion (algue unicellulaire) : deux cellules s’accolent (en position perpendiculaire) par leur mucilage. Les contenus cellulaires vont fusionner pour former un zygote.
L’hologamie chez Spirogyra. Deux filaments vont fusionner. Tout passe dans un des filaments. Il y a anisogamie car on trouve un donneur et un receveur.
2- Illustrations de quelques cycles.
a- Ulva lactuca.
On trouve deux générations (une haploïde et une diploïde), c’est un cycle digénétique haplodiplophasique. Les générations haploïdes et diploïdes ont la même morphologie (cycle digénétique isomorphe). Il y a planogamie anisogame.
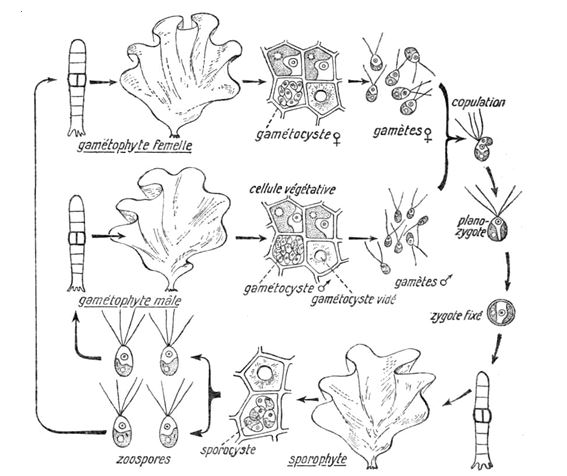
b- Codium sp.
Le cycle est monogénétique, diplophasique. La planogamie est anisogame. Les utricules portent des gamétocystes qui donnent des gamètes mâles et femelles qui après fécondation donnent un embryon qui va donner des filaments puis une plante qui porte des utricules.
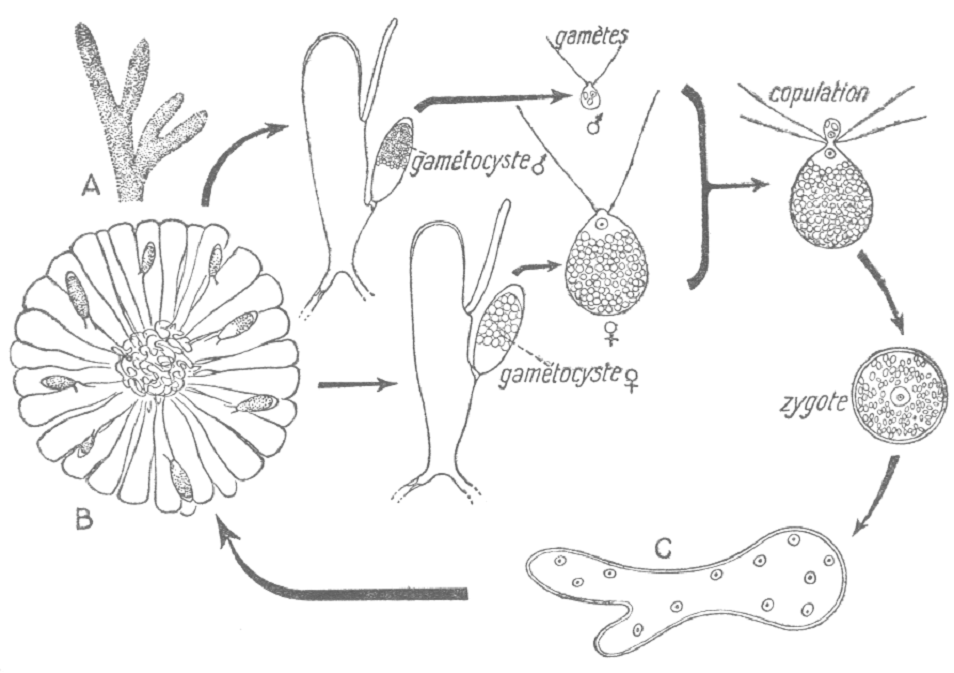
c- Chara sp
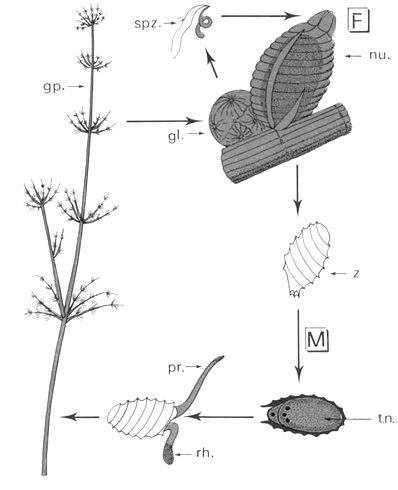
Le Chara a un cycle monogénétique et est haplodiplophasique. La fécondation est une oogamie.
La plante présente des gamétocystes mâles (ronds) qui vont libérer par éclatement, des spermatozoïdes biflagellés. Ces spermatozoïdes vont féconder l’oosphère qui reste protégée par une cellule spirale (àgamétange). Il va y avoir réduction chromatique qui aboutit à une cellule à un noyau et à une cellule à trois noyaux. La cellule mononucléée donnera la cellule germinative alors que la cellule trinucléée donnera les réserves en amidon.
==> Chara pourrait être le chaînon manquant entre les algues et les végétaux supérieurs. « L’œuf » de Chara préfigure la graine car il y a apparition des cellules de protection, d’albumen et d’une cellule germinative.
d- Ectocarpus siliculosus.
Ectocarpus présente deux cycles : un cycle digénétique diplohaplophasique et une variante monogénétique diplophasique.
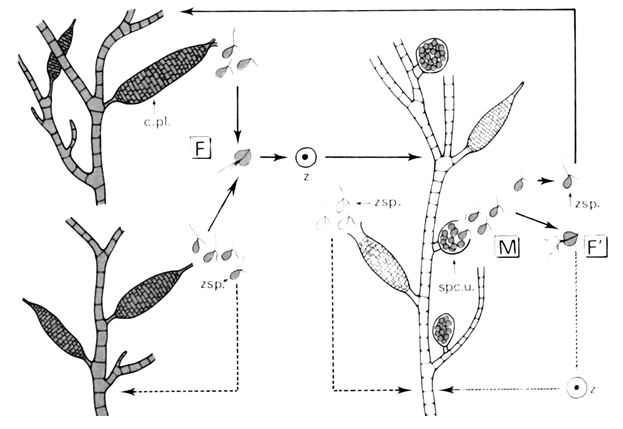
Les gamétophytes haploïdes portent des cystes pluriloculaires qui libèrent des zoospores. Ces zoospores ont deux devenirs possibles : soit, donner un gamétophyte haploïde par germination, soit, participer à la fécondation par planogamie anisogame.
Cette fécondation donne un embryon qui va former un sporophyte diploïde. Sur ce sporophyte, on va trouver deux sortes de cystes.
Les premiers cystes sont les cystes pluriloculaires qui donnent des zoospores à 2N. Ces dernières redonneront directement des sporophytes diploïdes. Les seconds cystes sont les cystes uniloculaires qui libèrent des zoospores à N. Ces zoospores vont alors donner un gamétophyte haploïde. Là, le cycle est bouclé.
On peut avoir une variante ou les zoospores à N jouent le rôle de gamètes et permettent donc la fécondation. Celle-ci donne un embryon évoluant en sporophyte qui pourra redonner des zoospores à N.
e- Fucus serratus.
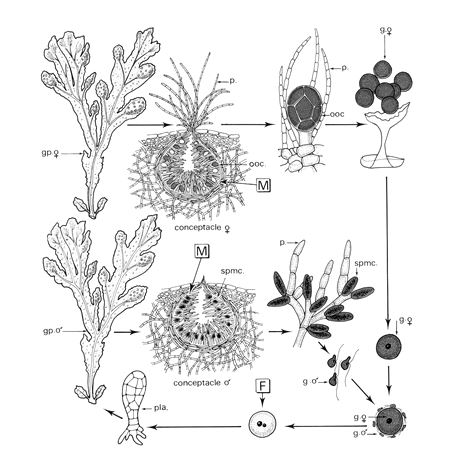
Le cycle de ce Fucus monogénétique, diploïde. La fécondation est une oogamie et les flagelles portés sont au nombre de deux et différents morphologiquement.
f- Les Laminaires.
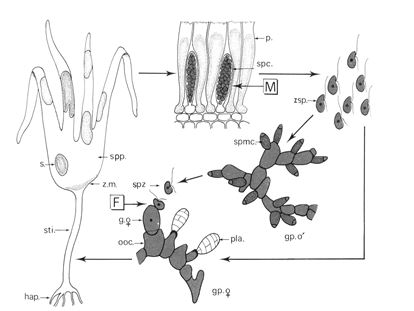
Le cycle présente deux générations, une génération sporophytique et une génération gamétophytique.
Le sporophyte va donner une cellule qui va subir la méiose, donner des cystes uniloculaires. Ces cystes sont insérés entre des cellules stériles, les paraphyses.
Les cystes contiennent des zoospores haploïdes qui donneront des gamétophytes (mâles ou femelles). Ces derniers (les formes mâles) forment des spermatocystes libérant des spermatozoïdes qui vont féconder les oosphères et ainsi, donner un zygote.
==> Le cycle est digénétique, haplodiplophasique, hétéromorphe.
g- Porphyra umbilicalis – algues rouges.
L’appareil végétatif est un cénobe.
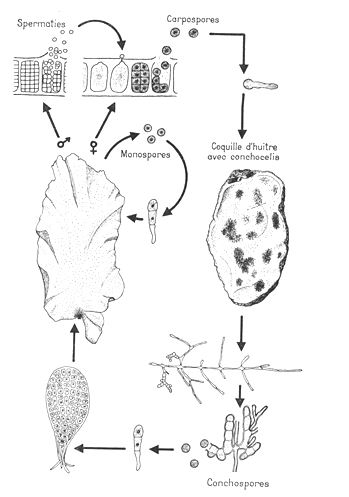
Il n’existe pas de cellules flagellées chez ces algues. On ne trouve donc pas de spermatozoïde mais des spermaties. Ces dernières vont féconder le carpogone qui subit alors trois mitoses et forme huit cellules. Ces huit cellules libérées sont les carpospores.
Elles vont germer et donner un filament incrustant (sporophyte qui se développe sur un support calcaire). Le filament subit la réduction chromatique et donne des spores qui en germant, donneront des lames de Porphyra.
h– Polysiphonia elongata (Florideophyceae).
Polysiphonia présente un cycle trigénétique, haplodiplophasique.
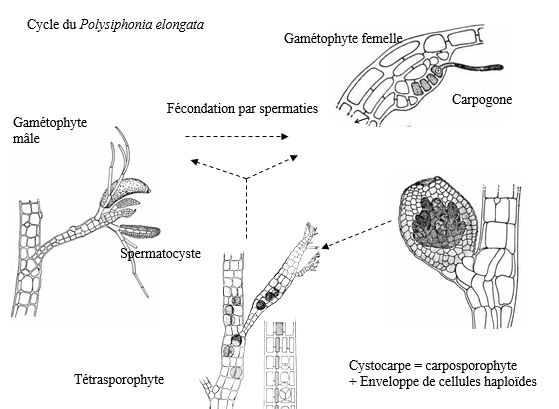
Le gamétophyte mâle libère des spermaties. Les gamétophytes femelles mettent en place des carpogones, surmontés par un trichogyne. Les spermaties vont féconder les carpogones.
La fécondation donne un zygote qui reste fixé sur le pied mère pour effectuer une multiplication cellulaire : il évolue ainsi en carposporophyte. Ce dernier libèrera des carpospores évoluant en (tétra)sporophytes lui-même, donnant des tétrasporocystes, libérant des tétraspores. Ces dernières spores donneront les gamétophytes (mâles et femelles).
Une génération est une structure pluricellulaire venant d’une fécondation ou de la germination d’une spore et qui libère elle-même des spores.
IV- Les algues et leur milieu.
A- Les types trophiques.
Les milieux de développement des algues sont des zones avec présence de lumière. Le classement se fait selon les sources d’énergie et selon les sources alimentaires.
Les bactéries vertes seront trouvées dans les sources d’eau chaude alors que les bactéries pourpres sont présentes dans les zones soufrées.
On pourra distinguer des organismes à photosynthèse archaïque ou à photosynthèse évoluée.
==> On fera une différenciation par rapport à la composition pigmentaire.
La bactériorhodopsine est une protéine permettant de faire toute la chaîne de la photosynthèse à elle seule.
Les plantes effectuant une photosynthèse évoluée comprennent les pigments suivants : fucoxanthine, chlorophylle A, photosystèmes, pigments surnuméraires comme les phycobilines (phycocyanine, phycoérythrine).
Les phycobilines sont surtout présentes chez les cyanobactéries et les rhodophytes.
- Le trajet des électrons.
Chez les algues et les cyanobactéries, le principe est le même : l’eau donne l’oxygène qui sera libéré sous forme d’O2 gazeux. Les électrons seront ensuite déplacés.
Les bactéries pourpres et vertes ont un photosystème anoxygénique qui montre un trajet cyclique des électrons.
B- Les algues benthiques.
1- Le milieu marin.
La flore marine est composée à 93,8% d’algues ; 5,5% sont des monocotylédones marins (exemples : Zostères, Posidonie) ; quelques lichens dans les zones de balancement de marées.
2- Les contraintes à la vie marine et adaptations.
On assiste à une raréfaction de la lumière en fonction de la profondeur. Plus la profondeur augmente, plus la quantité et la qualité de la lumière diminuent. Le rouge et le violet disparaissent les premiers. Le vert reste fort longtemps.
On observe donc :
- Des phytoplanctons dans les premiers mètres qui absorbent surtout le rouge et le bleu.
- L’énergie photonique est inversement proportionnelle à la longueur d’onde. (E=h.c/l)
a- L’adaptation aux profondeurs.
- La théorie de l’adaptation chromatique complémentaire.
En surface, toutes les algues sont viables. Un peu en dessous (à partir de 10 mètres), les algues vertes meurent. Les radiations vertes restantes ne permettent que le développement des algues rouges. Cette théorie est sûrement vraie pour les zones pauvres en espèces.
- la théorie de la plasticité de l’appareil pigmentaire.
Pour cette théorie, il faut faire une étude du rendement synthétique.
Avant un certain point d’éclairement, la concentration en oxygène est inférieure à 0, il y a donc respiration.
Au dessus, la concentration en oxygène devient supérieure à 0, il y a donc plus d’O2 dégagé que d’O2 utilisé pour la respiration.
Si la lumière diminue, le point de compensation va diminuer.
Si le point de compensation diminue, cela permet à l’algue un développement dans une illumination plus faible.
En fait, seul le rapport de la chlorophylle A sur la chlorophylle B augmente. Il y a donc augmentation de la taille des antennes collectrices de photons.
==> Chaque espèce s’adapte différemment.
b- Les contraintes liées à la vie en zone côtière.
Les marées vont provoquer une émersion des algues : il va y avoir une forte variation de l’illumination et une variation de pH et de salinité (on comprend alors le rôle de la paroi).
Les alginates sont des sucres acides avec des fonctions carboxyles qui piègent les ions en limitant l’entrée et la sortie d’ions au travers de la paroi.
L’illumination directe va entraîner la formation de radicaux libres (H2O2) qui entraînent une photo inhibition. Cette dernière est souvent irréversible.
Certaines algues vont développer un système de protection : présence d’un xanthophylle (violaxanthine) qui est transformé en zéaxanthine. La violaxanthine va modifier l’absorption car la zéaxanthine a un spectre d’absorption très fortement semblable à celui de la chlorophylle. Ce système est réversible quand la plante repasse à une plus basse illumination.
3- Zonation des algues.
Dans les zones de balancement de marées, il y a dispersion des algues selon les capacités de résistance aux contraintes liées à l’émersion. Les phéophycées sont spécialisées dans l’adaptation.
Pour plus de détails télécharger les documents ci-dessous:
Cours sur les Algues N°1
Cours sur les Algues N°2
Cours sur les Algues N°3
I liked it!!